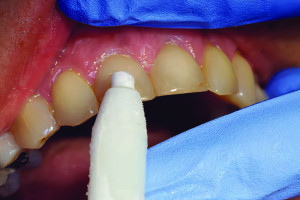
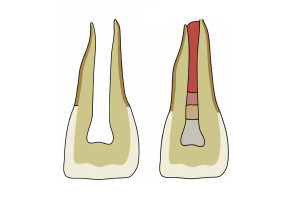
Хотя дентин и пульпа зрелых зубов различаются по составу и физическим свойствам, они имеют одинаковое происхождение в процессе развития и представляют собой единую функциональную единицу. Одонтобласты - клетки, образующие дентин, — находятся в пульпе на протяжении всей жизни зуба, а мезенхимальные стволовые клетки в пульпе могут дифференцироваться в дентинные одонтобласты. Таким образом, резидентные клетки пульпы, включая иммунные и стволовые клетки пульпы, а также одонтобласты, способствуют восстановлению дентина во взрослой ткани. Понимание молекулярных сигналов и биологических механизмов, которые управляют развитием соединения дентин-пульпа и приводят к дифференцировке одонтобластов с образованием продуктивного дентина, может способствовать разработке восстановительных методов лечения. В этой статье будет представлен обзор развития комплекса дентин-пульпа и представлены некоторые последние достижения в понимании молекулярных и клеточных механизмов развития дентин-пульпа и того, как они могут быть использованы в регенеративной стоматологии.
Формирование комплекса дентин–пульпа
Во время раннего эмбриогенеза клетки нервного гребня черепа нейроэпителиального происхождения расслаиваются и отделяются от нервной эктодермы, поскольку они претерпевают переход от эпителия к мезенхиме. Их новые мезенхимальные свойства, обусловленные эктодермальным происхождением, дали им уникальное название: эктомезенхима. Эктомезенхимальные клетки, происходящие из нервного гребня, вносят важный вклад в развитие головы и шеи, включая прорезывание зубов. Переход от эпителия к мезенхиме имеет решающее значение для развития зубов, поскольку эктомезенхимальные клетки, которые колонизируют нижнечелюстную и верхнечелюстную дуги первой глоточной дуги, являются предшественниками зуба и пульпы. Эктомезенхимальные клетки претерпевают морфологические и молекулярные изменения, а также приобретают мощный миграционный потенциал. По мере того как они мигрируют и заселяют глоточные дуги, эктомезенхимальные клетки пролиферируют и дифференцируются, реагируя на локальные сигналы в пределах своего миграционного пути. Реагирование на молекулярные сигналы, исходящие от соседней эктодермы полости рта, вызывает морфологические изменения, за которыми следуют молекулярные и функциональные изменения, превращающие клетки нервного гребня в черепе в зубной сосочек. Зубной сосочек будет еще больше дифференцироваться и даст начало пульпе зуба и окружающему минерализованному дентину (рис. 1). Поскольку только базальная мембрана отделяет зубной сосочек от вышележащей эктодермы, обе ткани остаются в тесном физическом контакте. Успешные взаимные эпителиально-мезенхимальные взаимодействия между эктодермой полости рта и слоями зубных сосочков обеспечат сигналы, необходимые для скоординированного развития этих двух эмбриональных слоев в зрелые ткани зуба, включая пульпу, дентин и эмаль (рис. 1).
Эти взаимные эпителиально-мезенхимальные сигналы инициируют молекулярные и клеточные изменения, которые стимулируют развитие зубов. Этот процесс распознается по ряду морфологических изменений, которые классически описываются как зубная пластинка, почка, колпачок, а также ранняя и поздняя колокольчатые стадии развития зуба (рис. 1). Другая, более поздняя классификация, которая подчеркивает функциональность, была использована для описания одонтогенеза в четыре фазы: инициация, морфогенез, дифференцировка клеток (цитодифференцировка) и расположение матрикса. Во время инициации эктодерма полости рта слегка утолщается в местах предполагаемых зубов, тем самым определяя зубную эктодерму, которая отмечает местоположение будущего соответствующего зуба, все еще в пределах континуума развивающихся дужек нижней и верхней челюсти. На стадии зубной пластинки эктодермальные клетки в этих будущих областях зубов подвергаются быстрой и асимметричной пролиферации, что приводит к их инвагинации в лежащую в основе эктомезенхиму, морфологически определяемую как зачаток зуба. Инвагинирующие клетки зубной пластинки подают сигнал соседним эктомезенхимальным клеткам, которые реагируют агрегацией, и передают сигнал обратно в зубную пластинку, которая расширяется, образуя трехмерный колпачок над агрегированной эктомезенхимой, морфологически распознаваемый как стадия колпачка. На стадии колпачка наблюдаются первые признаки гистодифференцировки, поскольку колпачок образует отчетливые слои. Самый внутренний слой накладывается на нижележащую эктомезенхиму и определяет ее как зубной сосочек вместе с охватывающим его зубным фолликулом, который будет способствовать формированию опорных тканей зуба, главным образом цемента и периодонтальной связки. Самый внутренний слой колпачка теперь дифференцировался во внутренний эмалевый эпителий и развивает жизненно важный сигнальный центр, отмечающий начало формирования коронки, известный как эмалевый узел. Сложные сигнальные пути внутри эмалевого узла и зубного сосочка и между ними, а также клетки внутреннего эмалевого эпителия направляют формирование бугра и, следовательно, влияют на морфологию коронки. Они будут описаны позже в этой статье. Недавние исследования выявили морфоген Sonic Hedgehog в качестве негативного регулятора образования бугорков. Замедленная съемка развивающихся зубных зачатков, помеченных флуоресцентными репортерами клеточного цикла, элегантно демонстрирует сложную координацию клеточной динамики и размножения, которые управляют формированием зубов.